Punctuated Equilibrium

Here, as in so much of the theorizing we have examined, there is a lot of confusion. The term itself, “punctuated equilibrium,” feels almost “made up,” and even scientists are divided about what the term really means.
But we are going to gain clarity in what follows. Punctuated Equilibrium (PE for short) was designed to address a particular problem in paleontology, and we will examine the problem and how PE is intended to solve that problem.
In 1971, biologist/paleontologist, Niles Eldredge, published a scholarly article, “The allopatric model and phylogeny in Paleozoic invertebrates,” which appeared in the journal Evolution. In this article, Eldredge followed the supposed evolution of eye lenses in a particular species of trilobite. He noted that the trilobites seemed to evolve in fits and starts, with relatively long periods of no change “punctuated” by relatively brief periods of change.
Now, please notice something important about this article.
Eldredge was not generalizing about what came to be known as the theory of PE. The article is about a very narrow topic and was not intended to address any particular general problem with the fossil record. Eldredge was narrowly focused upon one of the two major models of speciation: allopatric as opposed to sympatric speciation. The difference between the models is quite simple: allopatric speciation is when a speciation event results from geographic isolation, and sympatric speciation is when a speciation event takes place in situ within an otherwise homogeneous population. Just think about it this way: Speciation is either due to spatial separation between populations or not due to spatial separation. Eldredge was arguing strictly that allopatric (geographically isolated) speciation could happen quickly, due to the fact that the adapting population becomes geographically isolated from the “parent” population, allowing the changes to accumulate rapidly without ongoing genetic recombination with the “parent” population.
However, paleontologist, Stephen J. Gould, noticed that Eldredge’s specific point could be generalized to explain a universally-known problem in paleontology, which is that the “gaps” in the fossil record are worse than merely “gaps.” Even granting evolutionists their idea that the fossil record is a record of evolution, what the fossil record actually displays is long, long periods in which nothing is happening, “punctuated” by short periods in which “big leaps” take place. But nature doesn’t make “leaps,” and neo-Darwinism is fundamentally a theory of tiny-step-wise gradualism. So, the fossil record seems to be at least a serious anomaly for neo-Darwinism, if not an outright falsification of it.
Here is an image that clearly differentiates what neo-Darwinism would predict compared to what the fossil record actually shows.
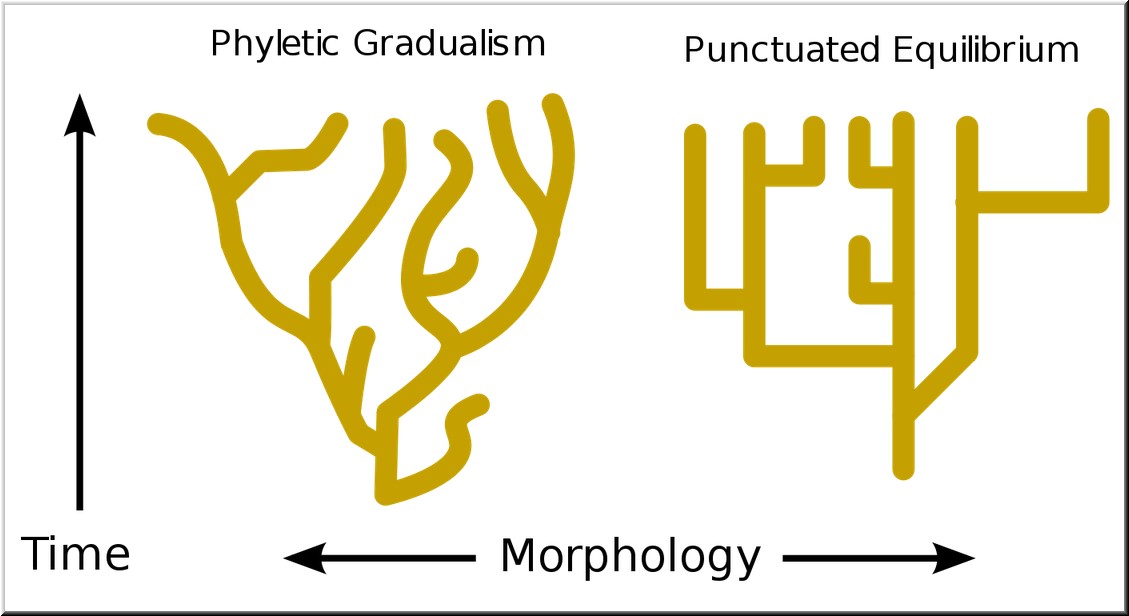 Smooth transitions vs. sudden appearances
Smooth transitions vs. sudden appearances
For the fossil record to be a record of gradualism, we would need to have discovered countless fossils showing tiny, step-wise changes over time. Instead, what the fossil record actually shows is long periods of time with no changes in species, and those long periods are “punctuated” by quick bursts of rapid (even sudden) change. Indeed, the fossil record indicates that “new species” almost always suddenly appear in the strata, with no “transitional forms” in evidence. And as we noted last week, Gould himself has repeatedly reported this general problem: “The extreme rarity of transitional forms in the fossil record persists as the trade secret of paleontology.”
If gradualism is true, the fossil record isn’t evidence of it! So, Gould early recognized the value in a theoretical response to the actual evidence of the fossil record. And he saw in Eldredge’s research a principle that could be generalized to account for the apparent “leaps” evidenced by the fossil record.
In 1972, Gould and Eldredge collectively wrote a book chapter as part of an anthology called Models in Paleobiology, entitled “Punctuated equilibria: an alternative to phyletic gradualism,” that described the general applicability of PE to the known problem in the fossil record. And in 2007, Gould published an entire book devoted to PE, entitled, fittingly enough: Punctuated Equilibrium, published by Harvard University Press. And between 1972 and 2007, Gould repeatedly refers to and refines PE as a theory that can explain the “gaps” in the fossil record.
To understand what Gould and Eldredge have really been arguing, and to see what the generalized theory theory of PE is, we must first clarify a terminological distinction.
Allopatric vs. Sympatric Speciation
One of the big questions in evolutionary biology is how significant the role of geographical isolation is. The allopatric vs. sympatric models of speciation are heavily studied exactly because the environment is held by Darwinism to play such a profound role in “adaptation” and hence speciation. So, evolutionists want to understand exactly how important geographic isolation is in the speciation process.
As we discussed the so-called “ring species,” we noted the profound role a geographical feature presumably played in the supposed “adaptations” that supposedly “accumulated” as an initial population migrated, split, encountered different environmental “pressures,” and ultimately experienced a “speciation event.” The geographic isolation (and, thus, the widely different environmental pressures) driving “ring speciation” is a classic fit with the allopatric speciation model.
Just as “ring speciation” is purported to occur very quickly, it is widely believed that allopatric speciation can (and usually does) occur much more quickly than sympatric speciation. Obviously, if a population becomes geographically isolated from its “parent” population, it will necessarily encounter different environmental “pressures” to which it will have to adapt, and these adaptations will accumulate more quickly simply because the “speciating” population is no longer interbreeding with the “parent” population. So the genetic and morphological differences will quickly accumulate, not “diluted” by constant interbreeding with the non-changing “parent” population, leading to a full “split” from the parent population until the “daughter population” can legitimately be considered a new species. Indeed many scientists believe that all forms of sympatric speciation actually still involve geographic elements on a small scale, such that there is no strictly sympatric speciation.
So, allopatric speciation is huge for neo-Darwinism exactly because that speciation model so neatly fits with the core idea of environmental pressures causing adaptations that accumulate to form new species. Only in the allopatric model of speciation is there an explanation of exactly why the changes accumulate in one part of a population and not another, thus causing one part to “branch off” into a new species, while the original population remains virtually unchanged.
Now, notice that the allopatric model of speciation is actually “orthodox” neo-Darwinism. It is still a speciation model within the confines of gradualism. Allopatric speciation is not suggesting that “nature takes leaps.” It is simply stating that evolution can occur must “faster” than usual, given the right conditions.
Thus, when we “contrast” the neo-Darwinian “orthodoxy” with the Gould/Eldredge theory of PE, we do so tongue-in-cheek to make this very point: PE just is “orthodox” neo-Darwinism! It is simply a general account of allopatric speciation intended to explain what we actually do observe in the fossil record. Scientists that read Gould/Eldredge to have “broken faith” with Darwinism are very confused about PE, and Gould himself suffered significant distress regarding how both the creationist camp and his own colleagues misunderstood his efforts. So, let’s be clear on this point: PE just is neo-Darwinism, and the allopatric model of speciation is now regarded by most biologists as the most fruitful model in which to study evolution.
To summarize the difference in the speciation models: allopatric speciation takes place as a result of geographic isolation and can (usually does) occur quickly because the “daughter” population’s changes are not being “diluted” with genetic material from the “parent” population; sympatric speciation takes place without geographic isolation, so the “daughter” population continues to interbreed with the “parent” population until (typically via the ecological species concept, ESC) the “daughter” population branches off into a new species, which obviously takes much longer and would leave behind a more “gradualistic” fossil record. Under both models, evolution is a gradual, step-wise transition between “forms” of creatures.
Why, Then, the Gaps?
PE explains the gaps as what we would expect to find in the fossil record! The theory, in summary goes like this:
* “Edges” of a population forage into new territory, where they encounter new environmental pressures.
* This small, even tiny, subset of the “parent” population begins adapting to the new environmental conditions.
* These adaptations are tiny and gradual. And the evolving population is very tiny. So the odds of any of these step-wise morphological changes being captured in the fossil record are infinitesimally tiny. In fact, the fossil record shows that these tiny changes in the tiny population are basically never “captured.”
* The fossil record will instead “capture” corpses from the large, unchanging “parent” population.
* Meanwhile, during long periods of time, evolution continues apace in the “fringe” population, which finally becomes reproductively isolated from the “parent” population.
* As complete reproductive isolation occurs, in the context of the ever-increasing geographical isolation, evolution becomes much more rapid.
* “Suddenly” (from the perspective of geological time), significant morphological changes are occurring in the “daughter” species, and that population is also growing. Meanwhile, the “parent” population is essentially unchanged. Now it can properly be said that there are two species.
* Corpses from the “daughter” species occur with high enough frequency that members of the new species start getting fossilized.
* Thus, the fossil record shows a long period of “stasis,” while evolution was quickly happening in the “daughter” species that just didn’t have enough members for them to be fossilized.
* The “punctuations” in the fossil record appear “sudden” only because it is a relatively “sudden” event for the “daughter” species to fully isolate itself and grow in population size sufficient to have members fossilized.
* But traditional, neo-Darwinian gradualism was happening all along, “sped up” in very localized fashion by the “fringes” of the original “parent” population.
This process can be illustrated by the above picture, now modified below to explain the “punctuations” and “stasis.”
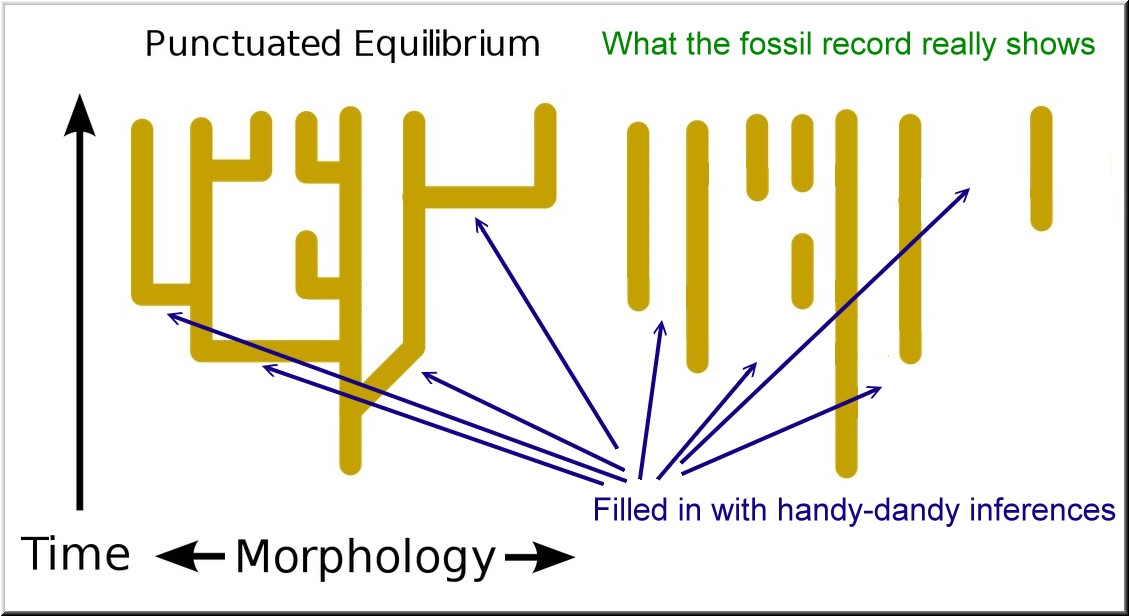 The “gaps” need an explanation.
The “gaps” need an explanation.
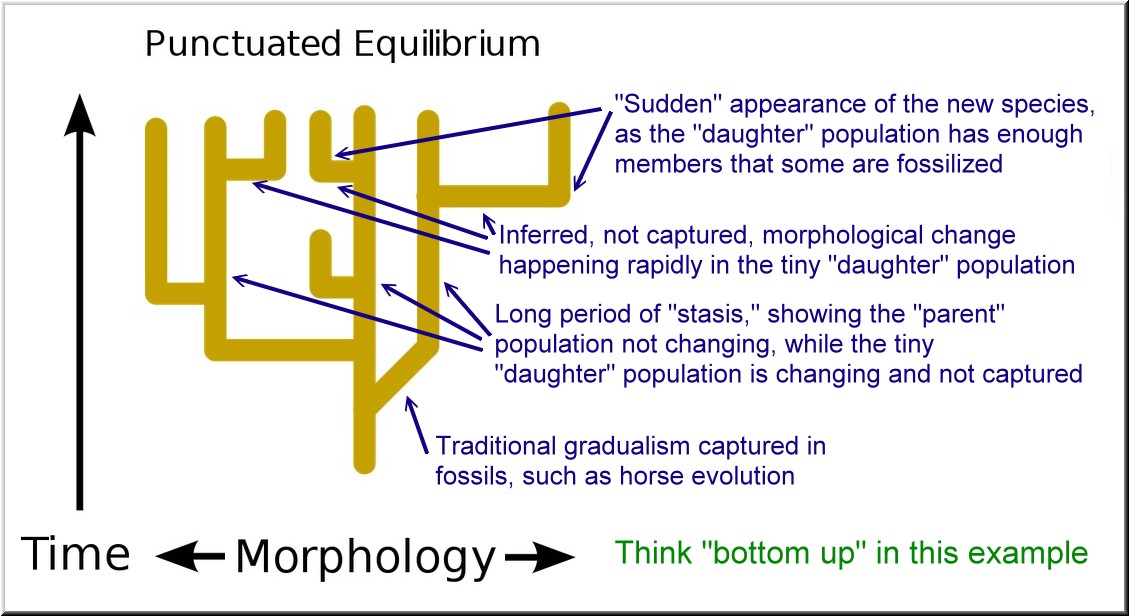 PE fills the “gaps” with a theory
PE fills the “gaps” with a theory
The horizontal lines in the above images are the brief periods of theorized “rapid adaptation and modification” that supposedly take place in the budding “daughter” population, while the “parent” population is in “stasis.”
The fabulous thing about PE is that it is a “gap theory!” It is, literally, a theory whose “evidence” is the actual lack of evidence! Because evolution is presumed to have occurred, then of course evolution must have been happening, even though there is zero evidence that it was. In fact, the theory actually states that evolution was happening because it was leaving no evidence.
Talk about no loose ends! That is about as tight (and circular) as “reasoning” can possibly be!
What “Ring Speciation” Tells Us
So, before any reasonable person could use PE as an explanatory approach to the fossil record, we would first have to have a reason to believe in allopatric speciation in the first place. Otherwise, as just noted above, PE would be a viciously circular argument and grand appeal to ignorance fallacy!
An appeal to ignorance fallacy is whenever you argue from some claim of ignorance to some substantive claim on that subject. For example, “There are no speed limit signs, and I don’t know what the speed limit is. Therefore there is no speed limit, and I can drive as fast as I want.” That example is obviously fallacious! However, appeals to ignorance can be more subtle….
1) Despite decades of searching, we have never discovered any Sasquatch corpses.
———————–
2) The population of Sasquatches must be quite small, so their corpses decompose before anybody discovers them.
Notice that the above argument must presume the existence of Sasquatches before it even makes any sense to “theorize” about why there is no solid evidence of their existence! After all, what would prove the existence of Sasquatches would be to “capture” either a live or dead actual Sasquatch! No such proof has been forthcoming, and one would expect the many Sasquatch hunters to have discovered some actual Sasquatch remains by now (decades of concerted searching). So, it is entirely reasonable to be skeptical of the existence of Sasquatches! But the above argument presumes their existence to explain why we remain ignorant of their existence. A way to recast the above argument shows what it is really claiming:
1) We have no positive proof that Sasquatches exist.
———————
2) This lack of proof is just what we would expect from a small, secretive population of Sasquatches.
Now it is clear that the very lack of evidence is being used as evidence, and this is typical of appeal to ignorance arguments.
So, what do neo-Darwinists need in order for PE to not be a grand appeal to ignorance fallacy? Well, in short, they need positive evidence indicating that allopatric speciation does in fact occur. With such evidence in hand, they could then go on to extrapolate that the fossil record is best explained by allopatric speciation, just as Gould/Eldredge theorized. That, at least, would not be an appeal to ignorance.
Scientists well-recognize that they need evidence of allopatric speciation. What most of them don’t recognize is that they do not have it. As we have seen, Irwin and other leading researchers recognize that we have never observed a speciation event, allopatric or otherwise. And the failure of “ring species” to evidence speciation, allopatric or otherwise, is actually a huge blow to the idea that we have good reason to think that allopatric speciation ever occurs! Yet most evolutionists believe that “speciation is happening all around us.”
If there is anything we learn from so-called “ring species,” it is that we actually have no evidence that speciation occurs, even in the supposed “classic” and “ideal” examples of it. So, we in fact have no solid evidence that allopatric speciation does occur. All of the “evidence” concerns “adaptations” that are then extrapolated (by presuming that speciation can occur) into an idea of “accumulated adaptations” sufficient to produce enough genetic variation as to produce a biological species concept (BSC) new species. But we have never observed anything of the sort, and we actually have solid evidence that certain genetic boundaries cannot be crossed.
No Evidence Of Speciation
Leading scientists readily admit that we have never observed a speciation event in our time, “because speciation occurs over long periods of time, much longer than even some generations of human observation.” So, the best evidence scientists have that speciation has occurred (which grounds their belief that they are studying “evolution” when they study adaptation) is inferred from the fossil record.
We will not appeal to ignorance ourselves, such as saying, “We have never seen a speciation event first-hand,” as though that is positive evidence against the possibility of speciation. Instead, we will look at the necessary inferences of paleontology to show that these cannot in principle be evidence for speciation. We will then simply state that we don’t believe in speciation, just as we (I hope) don’t believe in Sasquatch, because we have no solid evidence to suggest that there is any such thing. It is not an appeal to ignorance fallacy to argue from an ignorance claim to an “I’m not convinced” claim. The following inference is always legitimate:
1) I have no solid evidence to suggest that __________ exists.
—————–
2) I am not convinced that __________ exists.
So, let’s look at the fundamental inference of paleontology and see if it can withstand scrutiny.
Evolutionists love to trot out fossil evidence of horse evolution. As we argued last week regarding whale evolution, the fossil evidence is fragmented, partial, and highly interpreted. We actually have no solid reason to think that any of the “pre-horse” fossils have anything to do with modern horses.
By contrast, what we actually observe is that you cannot take even two horse-like creatures (horses and donkeys) and produce anything but an “evolutionary dead end” with them: mules, which are almost always sterile and which cannot produce viable offspring with any other creature. Apparently, the relatively minor morphological differences between horses and donkeys belie significant genetic differences, such that “underneath” their apparent “similarity,” horses and donkeys are actually dissimilar enough that their offspring “have nowhere to go” in an evolutionary sense. So far from horses and donkeys providing any evidence that morphologically “similar” creatures are really reproductively similar, we instead see that the genetic boundary between even such morphologically “similar” creatures is actually a deep divide.
Now, of course, evolutionists could turn this evidence on its head and claim: “See! Just as we would predict, horses and donkeys have already evolved to the point that they are properly different species. Now they cannot successful interbreed, so we see that a BSC event has occurred. We are seeing the fossil record of horse evolution continuing before our very eyes!”
But here’s the problem that evolutionists have with this sort of interpretation of the fossil record. We see huge morphological differences between breeds of dogs and between breeds of cats. In fact, the differences are so dramatic that if a “sequence” from Dachshund to Saint Bernard were discovered in the fossil record, it would be hailed as the “best evidence yet discovered, showing a gradualist, unbroken sequence of dog evolution.” After all, this “sequence” would be far more impressive than the supposed whale evolution, with claims about spine length, leg length, muzzle length, cant of the pelvis, and so on… all bone-structure inferences to “indicate” how Dachshunds were “transitional” from “earlier forms” and then gradually morphed into the large, stately Saint Bernards that we know as “dogs” today. If there were such a “record” of dog evolution, it would only be stymied by the fact that we see all of these “transitional forms” of dogs today, so we know that the supposed “sequence” really reflects breeds rather than species. So, huge morphological differences do not translate into reliable inferences about speciation!
Conversely, really, really tiny morphological differences do not translate into reliable inferences about “similarity” from a BSC standpoint, as we know from horses and donkeys! Vast morphological differences between breeds of dogs do not indicate different species. And tiny morphological differences between horses and donkeys do indicate genuine reproductive isolation. So, morphology cannot be mapped onto speciation, yet that is precisely what paleontology must do! And that is the fundamental problem that paleontology has as a science. Its most fundamental working principle is demonstrably false. From morphology, you cannot reliably infer “similarity” or speciation!
And paleontology must make this fundamental inference on the basis of the presumption that genetic boundaries are quite fluid and permeable. Yet, evolution in general also needs these permeable genetic boundaries to ultimately result in non-permeable genetic boundaries! After all, there must be smooth, gradual “gene flow” right up until the moment of a speciation event. And there, the gene flow must truly “branch” to ensure that genuine “kind” boundaries do form, because we know that hard “kind” boundaries do exist (e.g.: humans and elephants).
Horses and donkeys show that there are genuine genetic boundaries between even morphologically very similar “forms.” Meanwhile, endless experiments with bacteria, fruit flies, mice, and other fast-breeding creatures all show that even significant genetic modification does not get the “kind” across the BSC boundary to new species.
So, paleontology is at best a “soft science” that relies so heavily upon a demonstrably false fundamental principle that its inferences are perpetually suspect (and commonly overturned by paleontologists themselves).
Perhaps you will remember from some weeks ago when we talked about empirical observables and “unobservables in principle.” We said that observables are those entities or processes that we directly observe with our senses. I can directly observe that I have hands. The very things that I call “hands” are the self-same things that I directly observe with my senses. And, regarding processes, I can directly observe the motion of water down a stream and thus correctly report, “The stream is flowing.”
But we often use both nouns and verbs to refer to entities and processes that we cannot accurately report as we observe them. When a chemist, for example, describes what is the “makeup” of my hands, she is using terms to describe entities and processes that I cannot in principle directly observe. Instead, I “observe” my “hands” in that chemical sense only through a microscope. There is a layer of inference in such an “observation” because I now have to build in a theory about light refraction, wavelengths of light, and so on.
At the far end of the “observable” continuum are purported entities and processes that can never in principle be observed via the senses. For example, we cannot in principle observe both the position and velocity of an electron at the same time. We cannot in principle observe quarks with our senses. We can only employ inferences based upon the readings of instruments. What we are sensing in the instruments is not the quarks. What we are sensing are the inferred effects of the quarks.
This is not a rigorous distinction by any means. But its intuitions are solid, and we actually employ these sorts of distinctions in the everyday world to determine reliability! In a court room, for example, an attorney might ask, “Did you actually observe the gun in the suspects hands, or did you just infer the gun from the way his hand was positioned?” We recognize a distinction between direct observation and inferences. And we can recognize when inferences are becoming further and further removed from direct observation as they become more and more heavily dependent upon inference instead of observation.
The more “purely inferential” a particular “observation” is of an entity or process, the less plausible it is that our “report” of our “observation” is accurate. And scientists often engage in some pretty strident talk about entities and processes that are unobservable in principle.
A classic example of unobservable-in-principle talk concerns the distant past, particularly the past in which no witnesses were in principle possible. Even the recent past is sketchy! And the further back you go, the less likely it is that your “historical research” is producing a truly accurate picture of entities and events! It is for this reason that history as a discipline (and the historians I have personally known) are quite humble and mitigated in their “assertions” about historical entities, events, and the interpretations they draw from their strongest evidences. By contrast, scientists are often downright arrogant in their pronouncements regarding entities and events supposed to have existed and occurred in the dimly distant past.
Paleontology is a soft science that almost invariably deals with fragmented, disassociated hard parts (bones, shells, teeth, etc.) of supposed creatures, and from these largely discontinuous fragments imagination and the “wish to believe” fills in the pieces (often adding entire tails, limbs, or even bringing parts of multiple animals together into one). From this “assembly” process, “creatures” emerge, replete with skin color, locomotion, habitat, dietary and mating habits, and, yes, species!
As we have seen, it is all “soft,” and even paleontologists themselves overturn what they once so stridently asserted. Far from typical historical humility, the train of assertions and recantations in paleontology is long. We have not even scratched the surface thus far!
So, the state of paleontology is this:
* Its inferences depend upon a morphology -> speciation principle that is demonstrably false.
* Its inferences work with precious little actual “hard parts,” depending instead upon filling in the missing pieces with unobservables-in-principle.
* Even what “hard parts” paleontology has to work with do not reliably “resurrect” the animal whose parts they were; so “the animal” as paleontology reconstructs it lack many and critical aspects of morphological analysis (soft parts, behavior, actual environment, etc. are all missing).
* Paleontology presumes that evolution has occurred, so all interpretations are deeply circular:
1) Actual speciation has never been directly observed.
2) Evolutionists point to the fossil record for evidence that speciation has occurred.
3) But the fossil record can only be interpreted as evidence of speciation if it is already established that speciation has occurred.
4) But (1) above.
5) So (2) above.
6) But (3) above.
7) Rinse and repeat as needed.
——————-
8) Voila! Evolution is a fact, with abundant evidence everywhere we look!
And the theory of PE takes this circularity one step further to add a full-blown appeal to ignorance fallacy:
1) We don’t know of “transitional forms” in the fossil record; there are indeed “gaps.”
2) If speciation was occurring, there should be countless “transitional forms” in the fossil record.
3) Because of the “gaps,” we know that speciation was occurring.
4) The “gaps” themselves demonstrate that PE was how speciation was occurring
——————
5) PE speciation is demonstrated by the lack of evidence that speciation was occurring.
Stupid Creationists
Believers in intelligent design, including full-blown young-earth creationists, are eviscerated in the intellectual community and in public as ignorant, stupid, blind-faith idiots, and dolts who are unwilling to see the evidence for what it really is!
In fact, biologists point to paleontology as the evidence of speciation that they are supposedly studying when they study “adaptation.” And paleontologists point to biology, where they presume that speciation events are well-documented, as the foundation of their fundamental inference of speciation in the fossil record.
It would be like a comedy, with two people standing outside a locked car, both saying, “I thought you had the keys!” But it’s no comedy. The circularity is deep and viciously fallacious.
If it were not for the stridency with which scientists assure the public that “evolution is a fact” and that the evidence of speciation is everywhere we look, they might be forgiven for simply believing and humbly sharing how it seems to them. But soft sciences such as paleontology try to borrow the credibility of hard sciences, such as physics. As a result, the soft scientists help themselves to a confidence (arrogance) that they are not entitled to have.
What it turns out is that human beings, trained observers or not, are subject to a psychological phenomenon called “inattentional blindness.” It is worth your time to conduct a bit of online research about this. Here is just one teaser.
Here is one of the videos I showed during lecture.
While the phenomenon is typically associated with visual perceptiveness, it can afflict anybody in any setting that demands close attention coupled with expectations of what they are looking for.
Of course, the scientific community is believed to be “self-correcting” insofar as a large number of scientists “looking” at the same evidence are unlikely to all be fooled. But here’s the irony in the paleontological community: The community often does correct the mistakes of researchers, yet members of the community then just proceed to make exactly the same sorts of mistakes regarding other data. Sometimes it’s the same researchers, and other times it’s other researchers. But the community is rife with flagrant error, then correction/recantation, then the same sorts of errors all over again. So, although “science is self-correcting” even in paleontology, it is not self-correcting in the way that most matters! Paleontologists are not asking themselves critical questions about what they are expecting to see and whether the evidence really points to what they expect to see or just in any direction anybody happens to prefer.
So, they focus their attention closely on minute details of a particular edge of bone, trying to see where the fragment fits into a “whole skeleton” without looking at the big picture of the “whole skeleton” or the bigger picture of whether or not this “creature” is anything like how their mind’s eye “sees” it. Worse, they never back up to ask the even bigger-picture questions about how to place such “creatures” in an overarching taxonomy. Oh, they “consider” that question, but they do so strictly in the “seeing what you expect” perspective of speciation.
Thus, really smart people that are highly trained and very observant fail miserably to see the flagrant inferential problems in their “assembly” of the “evidence.” And, thus, really smart people like Gould, Eldredge, and their surrounding community can “see” evolution in the gaps; they can “see” evidence of speciation from the actual lack of evidence.
Before we bash on them too hard, however, let’s remind ourselves again that we creationists have legitimately earned the ire and indictments directed at us. While we chuckle at how scientists can miss the “elephant in the room” in their own research, let us be meek and humble, because we have been (and are) profoundly guilty of elephants in our own rooms. This is the human condition, and I cannot cease to make the same point again and again: Intellectual Honesty! Intellectual Honesty!
Continually ask yourself what presumptions and inferences you are bringing to bear on your own interpretation of the evidence. Ask yourself what evidence you are ignoring because it doesn’t “neatly fit” within your present web of beliefs. Ask yourself whether you are primarily a comfort-seeker who prefers to never have that web jiggled in any way.
If we are going to hold scientists to the standard of solid interpretations and inferences from the evidence, then we must hold ourselves to that same standard. And we must be quick to “suffer rebuke” and recant badly-held perspectives.
Let us not give any cause to be legitimately called “Stupid creationists.”